
tft lcd loses cohesion made in china

China"s first 8.5-generation TFT-LCD production line was launched in Bengbu, East China"s Anhui province, on June 18, 2019, representing a breakthrough in the production of high-definition LCD screen, Science and Technology Daily reported.
TFT-LCD, or Thin Film Transistor Liquid Crystal Display, is key strategic material of the electronic information display industry. The Gen 8.5 TFT-LCD production line, launched by the Bengbu Glass Industry Design and Research Institute of the China National Building Material Group, will produce high-definition LCD screens of 55 inches, the report said.
According to the Liquid Crystal Branch of the China Optics and Optoelectronics Manufactures Association, the demand for TFT-LCD in the Chinese mainland was about 260 million square meters in 2018, including 233 million square meters" Gen 8.5 TFT-LCD. However, the annual supply of domestically made TFT-LCD is less than 40 million square meters, with all of them Gen 6 or below, which cannot meet the demand in scale and quantity.
The association predicted that China"s market demand for Gen 8.5 TFT-LCD or above will exceed 300 million square meters by 2020, accounting for 49.6 percent of the total global demand.
The production and control precision of Gen 8.5 TFT-LCD is comparable to that of the semiconductor industry, representing a higher level of large-scale manufacturing of modern glass industry.
The institute in Bengbu, with 60 years of expertise in glass, has finally made a breakthrough in the production of Gen 8.5 TFT-LCD, and will provide key raw material guarantee for China"s LCD panel industry after it goes into mass production in September, the report said.
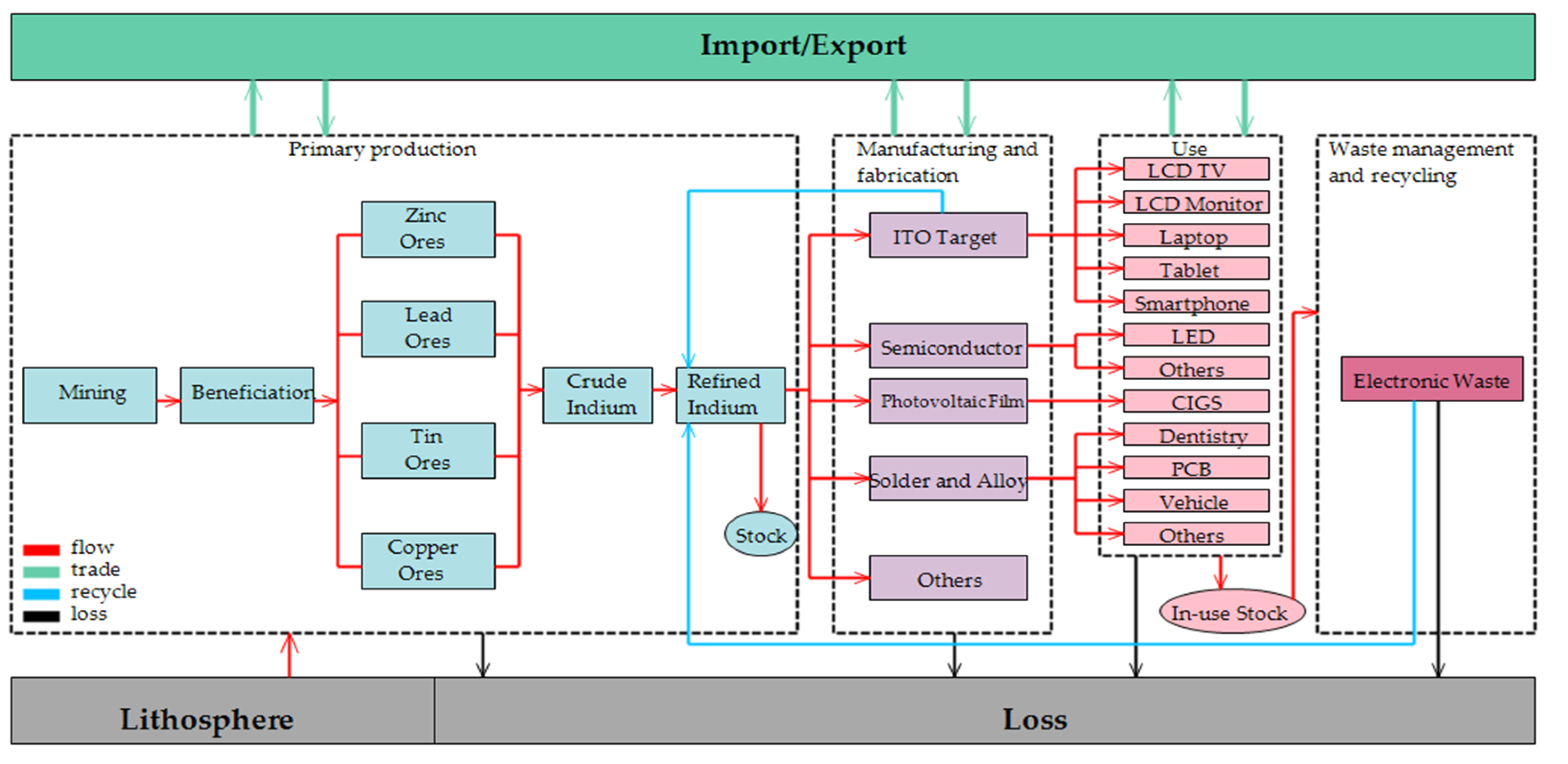
Washington must take note of these realities and invest in low-cost and specialized, skill-based workforce training for employees in vital industries, in addition to deepening its pool of advanced scientists and engineers. Bureaucratic reorganization could help focus government attention and cohesion on this problem: Consolidating the Department of Education and Department of Labor into a Department of Education and the Workforce would create a single entity dedicated to endowing Americans with the skills they need throughout the full continuum of their professional lives. And while it’s true that not all jobs will return, focusing training on high-tech manufacturing will prepare a workforce that attracts new, potentially higher-paying jobs to U.S. shores.

We have now more fully described the ChIP-seq data on MCM2. We did not observe many MCM2 peaks using MACS2, perhaps due to the rather broad localization of the MCM complex in early S phase cells. In the revised manuscript, we used a clustering approach called SICER to identify of MCM2 enriched domains. We could identify many such MCM2-enriched domains. When NIPBL (SCC2) peaks were used as the reference, these MCM2 domains and SCC1 peaks were found to co-localize with 1,041 NIPBL peaks. We did not perform ChIP-seq on CTCF. Homer de novo motif analysis revealed that the CTCF motif was the top DNA consensus sequence for RAD21 (SCC1) binding, with scores of 0.915 and 0.914 in two independent experiments. In addition, we can readily observe co-localization of RAD21 with CTCF in the human genome using the CTCF ChIP-seq data in the ENCODE database. These findings confirm that cohesin is enriched at CTCF sites. Because CTCF does not impair sister-chromatid cohesion, we reason that the pool of cohesin enriched at CTCF sites might be different from the pool that associates with NIPBL and MCM. These new analyses are included in Figure 2—figure supplement 1. The ChIP-seq data have been deposited into GEO (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE112028).
Subsection “Replisome components promote the establishment of sister-chromatid cohesion”, first paragraph, Figure 4A. In order to examine cohesion status, the authors performed a FISH assay using a probe that specifically recognized a locus on chromosome 3. No information was provided on the replication status of that locus. In fact, it is well-known that almost 50% of genome is replicated early in S phase and the remaining 50% is replicated late. FISH experiments should be performed using two different probes (replicated early and late) at least in both unsynchronized and synchronized cells.
WDHD1/TIMELESS, or RPA2 with the 465K16 probe that detects the late-replicating locus (Figure 6—figure supplement 2B, C). Likewise, depletion of FEN1 or LIG1 did not impair cohesion at the late-replicating locus recognized by the 465K16 probe. Additionally, we performed FISH experiments in asynchronized cells with both probes (Figure 6—figure supplement 3). We observed an increased number of cells that had completed replication in both genomic loci when Sororin, WDHD1/TIMELESS, or RPA2 was depleted in the absence of thymidine synchronization, consistent with the notion that depletion of these replication factors causes DNA damage and G2 delay under these conditions. For the 466L19 probe, only the cells with two pairs of FISH dots were analyzed, and the distances between paired FISH dots were quantified. For the 465K16 probe, only cells with two or three pairs of FISH dots were analyzed, and the distances between paired FISH dots were quantified. As expected, we observed cohesion defects in cells depleted of Sororin, WDHD1/TIMELESS, or RPA2, but not in cells depleted of FEN1 or LIG1 using both FISH probes. These new data with an additional FISH probe and in cells without synchronization are entirely consistent with and greatly strengthen our original conclusions.
3) "Initial experiments in yeast showed that inactivation of the Scc2/4 complex in cells enriched in early S phase by hydroxyurea did not cause cohesion defect,… (Lengronne 2006)." That study was concerned with cell viability rather than a specific cohesion defect. Scc2 is not required to retain cell viability during S phase, though based on recent results from the Nasmyth and Uhlmann labs, it seems clear that cells develop a cohesion defect if Scc2 lacks in S phase that is compatible with viability. This could be rephrased.
We have rephrased this statement to more accurately describe the findings of the Lengronne study. We note, however, that the Lengronne study did specifically examine cohesion status. In that paper (on p789), the authors stated that “We further confirmed that sister chromatid cohesion was intact in scc2-4 cells that traversed S phase at restrictive temperature (8% cells with separated GFP-marked URA3 loci in G2/M, compared to 7% after release at permissive temperature).”
Depletion of the MCM complex in human cells did not apparently affect the early S phase arrest exerted by thymidine. As shown in Figure 5A, cells depleted of MCM2 treated with thymidine had a very similar cell cycle profile as compared to control depleted cells treated with thymidine. In the absence of thymidine, cells depleted of MCM2 for 48 hours ultimately arrested at G2. Thus, the depletion of MCM2 is partial, and does not completely block DNA replication. As we discussed, this partial depletion allows DNA replication to proceed (albeit less efficiently), thus revealing cohesion defects at the improperly replicated loci. To minimize complications from cell cycle differences, we performed cohesin immunostaining and immunoprecipitation under thymidine treatment conditions, which arrested cells depleted of various replication factors in early S phase (see Figure 5A).
13) The epistasis analysis of cohesion establishment factors in Figure 4B is confounded by potentially partial protein depletion by RNAi. Note that Scc2 depletion gives a relatively modest cohesion defect, which would be expected to result in no cohesion at all. RNAi of multiple components often results in further reduced depletion efficiency. Any conclusions about synthetic interactions, or lack thereof, should therefore be made cautiously. This also applies to the rescue of cohesion defects by WAPL depletion (Figure 4—figure supplement 1).
14) The interesting observation that FEN1 and nucleosome assembly components have no detectable role in sister chromatid cohesion establishment is in line with results in budding yeast (Borges et al., 2013), which could be mentioned.
15) The authors suggest that RPA and other replisome components contribute to the MCM-SCC2/4-cohesin interaction indirectly through regulating DDK. This is an interesting result that goes towards the mechanism of cohesin-replisome interactions. To substantiate this point, it would be interesting to quantitatively compare CDC7 loss from MCM2 with cohesin loss from chromosomes and sister chromatid cohesion defects following depletion of different replisome components. Do these measures correlate and follow a quantitatively similar pattern?
This is a good point. Inactivation of different replisome components led to reduced MCM2 phosphorylation at S53, impaired cohesin loading and cohesion defects to varying degrees. However, the MCM2 S53 phosphorylation levels did not appear to quantitatively correlate with the degrees of cohesion defects. It is possible that DDK sites other than S53 are functionally important. Alternatively, RPA and other replisome components directly contribute to NIPBL-cohesin binding. We have discussed this point more thoroughly in the revised manuscript.



 Ms.Josey
Ms.Josey 
 Ms.Josey
Ms.Josey